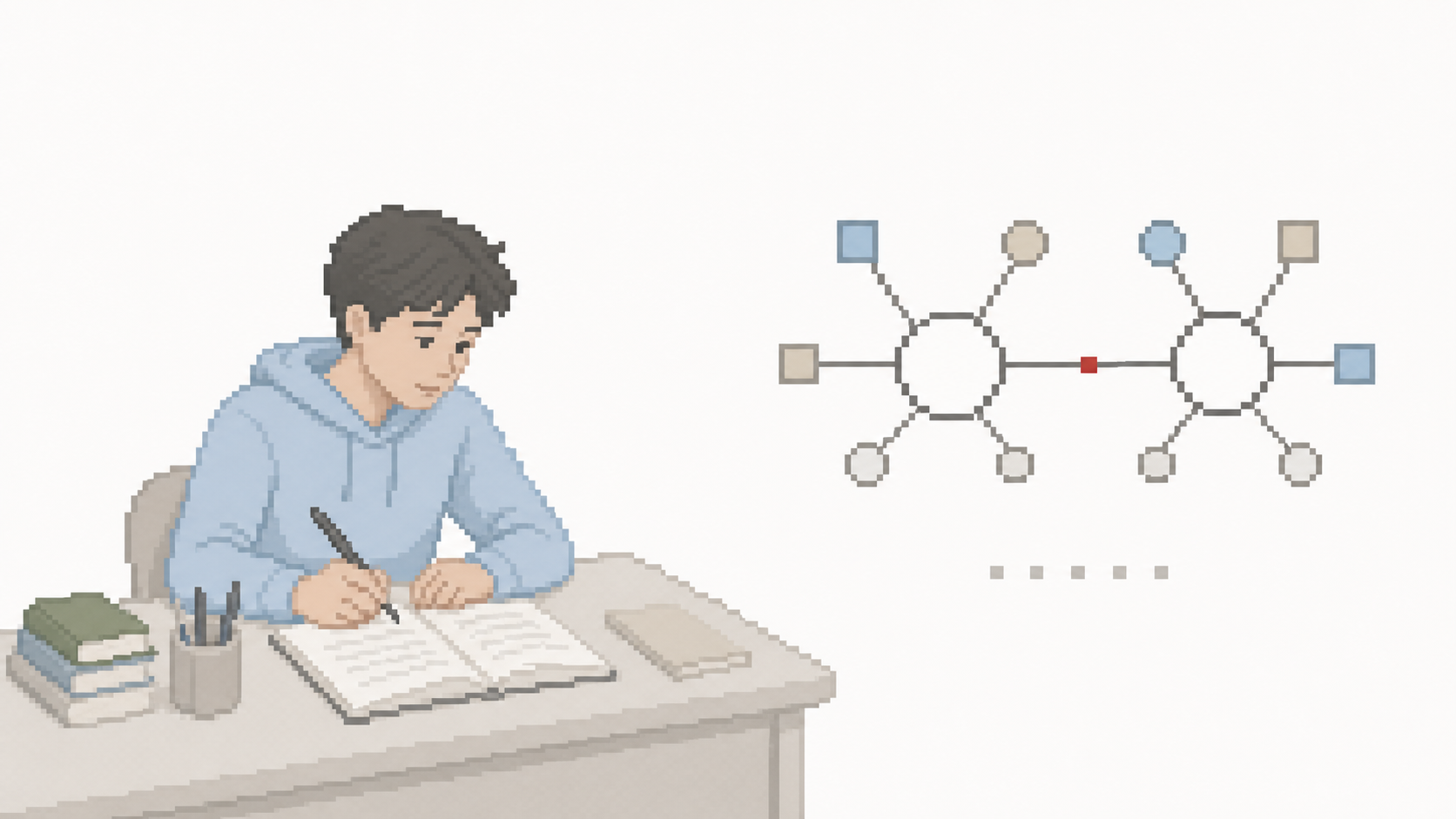
Si vous lisez un article scientifique sur les peptides, un manuel de biochimie, ou même une étiquette d'hydrolysat de collagène en parapharmacie, le terme *« liaison peptidique »* revient constamment. C'est le bloc de construction fondamental qui distingue un peptide (chaîne d'acides aminés liés) d'un simple mélange d'acides aminés libres.
Cette liaison covalente amide (–CO–NH–) entre le groupe carboxyle d'un acide aminé et le groupe amine du suivant a été caractérisée chimiquement par Emil Fischer entre 1899 et 1902 — travail qui lui a valu le Prix Nobel de chimie 1902 et qui a fondé la chimie peptidique moderne. C'est Fischer qui a forgé le terme *peptide* à partir du grec *peptos* (digéré) — référence à la formation des peptides lors de la digestion des protéines alimentaires.
Quatre angles : la chimie de la liaison (groupe amide, condensation, hydrolyse), la formation *in vivo* via les ribosomes lors de la traduction, l'hydrolyse inverse en digestion ou en hydrolyse industrielle, et l'importance structurelle de la planarité — conséquence géométrique qui rend possible la structure secondaire des protéines (hélices α, feuillets β).
1899-1902, Emil Fischer démontre la chaîne peptidique
Le contexte historique : à la fin du XIXe siècle, on sait que les protéines alimentaires sont digérées en composants plus petits absorbables, mais leur nature chimique précise reste débattue. Plusieurs hypothèses circulent : agrégats colloïdaux, composés de structure mal définie, mélange d'unités libres.
La démonstration de Fischer
Emil Fischer, professeur à Berlin, publie entre 1899 et 1907 une série de papiers où il :
- Caractérise chimiquement les acides aminés individuels
- Synthétise chimiquement les premiers dipeptides puis tripeptides en laboratoire — démontrant que la liaison amide est réversible et reproductible
- **Forge le terme *« peptide »*** à partir du grec *peptos* (digéré, qui peut être digéré)
- Propose la structure linéaire en chaîne d'acides aminés liés par des liaisons amides comme structure générique des protéines
C'est la première démonstration que les protéines ne sont pas des agrégats indéfinis mais des polymères linéaires d'acides aminés avec une séquence définie. Fischer obtient le Prix Nobel de chimie 1902, formellement pour ses travaux sur les sucres et les purines — la chimie peptidique poursuivie après le Nobel reste l'un de ses héritages majeurs.
La projection de Fischer
Au passage, Fischer a aussi introduit la projection de Fischer des sucres — un outil graphique de représentation stéréochimique encore largement utilisé en chimie organique aujourd'hui. La même approche s'applique aux acides aminés (configuration L des acides aminés protéinogéniques).
Mécanisme — condensation avec libération d'eau, hydrolyse en sens inverse
La formation d'une liaison peptidique entre deux acides aminés est une réaction de condensation :
H₂N–CHR₁–COOH + H₂N–CHR₂–COOH ↓ (avec catalyse, énergie) H₂N–CHR₁–CO–NH–CHR₂–COOH + H₂O
Le groupe carboxyle (–COOH) du premier acide aminé se combine au groupe amine (–NH₂) du second pour former la liaison –CO–NH– (amide). Une molécule d'eau est libérée dans le processus.
*In vivo* — les ribosomes
Dans la cellule vivante, chaque liaison peptidique d'une protéine est formée sur un ribosome lors de la traduction de l'ARN messager. Les acides aminés sont amenés par des ARN de transfert (ARNt), positionnés successivement dans le site catalytique du ribosome (sous-unité 50S chez les bactéries, sous-unité 60S chez les eucaryotes). La réaction est catalysée par l'activité peptidyl-transférase intrinsèque du ribosome — qui est une ribozyme (catalyseur basé sur l'ARN ribosomique, pas sur une protéine).
Un ribosome forme environ 15 à 20 liaisons peptidiques par seconde chez les bactéries en croissance — soit une protéine moyenne (~300 acides aminés) en 15 à 20 secondes.
*In vitro* — la synthèse peptidique solide
Pour la production industrielle des peptides médicaments (insuline, sémaglutide, octréotide, etc.), la formation des liaisons peptidiques se fait *in vitro* par la synthèse peptidique solide (Solid Phase Peptide Synthesis, SPPS (synthèse peptidique solide)) inventée par Bruce Merrifield en 1963 (Prix Nobel de chimie 1984). Chaque liaison est formée par activation chimique du carboxyle (souvent par un carbodiimide) puis condensation avec l'amine de l'acide aminé suivant. Voir synthèse peptidique solide pour le détail.
L'hydrolyse — réaction inverse
L'hydrolyse d'une liaison peptidique consomme une molécule d'eau et libère les deux acides aminés (ou peptides plus courts) :
H₂N–CHR₁–CO–NH–CHR₂–COOH + H₂O ↓ (catalysée par enzyme, ou acide/base + chaleur) H₂N–CHR₁–COOH + H₂N–CHR₂–COOH
*In vivo*, l'hydrolyse digestive est catalysée par des enzymes : pepsine dans l'estomac (à pH acide), trypsine et chymotrypsine dans l'intestin grêle, plus diverses peptidases pour finir le clivage en acides aminés libres absorbables.
Industriellement, l'hydrolyse contrôlée des protéines alimentaires (collagène bovin/marin, whey, soja, caséine) génère des hydrolysats peptidiques — peptides courts à activité biologique parfois utile (peptides ACE-inhibiteurs, peptides immunomodulateurs). Voir hydrolysat protéique.
Planarité de la liaison — conséquence structurelle
La caractéristique géométrique la plus importante de la liaison peptidique est sa planarité. Comprendre pourquoi elle est plane, c'est comprendre pourquoi les protéines ont des structures secondaires reconnaissables (hélices α, feuillets β).
Résonance partielle
Dans la liaison –CO–NH–, l'atome d'azote possède une paire d'électrons libres (paire non-liante) qu'il peut partager partiellement avec le carbone du carbonyle adjacent. Conséquence : la liaison C–N acquiert un caractère partiel de double liaison (~40 % selon les calculs de chimie quantique), tandis que la liaison C=O acquiert un caractère partiel de simple liaison.
Deux conséquences pratiques :
- Les six atomes impliqués (Cα-C-O-N-H-Cα suivant) sont contraints à être coplanaires — ils forment un plan rigide appelé *« plan peptidique »*.
- La rotation autour de la liaison C-N est restreinte — c'est cette restriction qui contraint la conformation 3D de toute la chaîne polypeptidique.
Les angles dièdres phi et psi
De part et d'autre du carbone alpha (Cα) d'un acide aminé, deux liaisons restent libres de rotation :
- L'angle phi (φ) entre N–Cα
- L'angle psi (ψ) entre Cα–C
Ces deux angles définissent à eux seuls la conformation 3D d'un acide aminé dans la chaîne. La combinaison des φ et ψ de tous les résidus d'une protéine définit sa structure 3D complète.
Le diagramme de Ramachandran
En 1963, Gopalasamudram Ramachandran propose un diagramme qui représente toutes les combinaisons possibles de φ et ψ pour un résidu peptidique. Ce diagramme révèle que seules certaines régions sont sterically accessibles — les angles dans ces régions correspondent précisément aux hélices α (φ ≈ -60°, ψ ≈ -45°), aux feuillets β (φ ≈ -120°, ψ ≈ 130°), et à quelques autres conformations.
C'est le travail théorique de Linus Pauling et Robert Corey en 1951 qui avait prédit ces structures secondaires avant leur démonstration expérimentale — sur la base unique de la planarité de la liaison peptidique et des règles d'encombrement stérique.
En pratique
Quand vous voyez un peptide thérapeutique (insuline, sémaglutide, octréotide) représenté sous forme de structure 3D, chaque liaison peptidique est un plan rigide. Les seuls degrés de liberté sont les angles φ et ψ aux jonctions. C'est cette rigidité partielle qui rend les peptides programmables structurellement — on peut concevoir une séquence pour qu'elle se replie d'une certaine manière, à condition de respecter les contraintes Ramachandran.
La liaison peptidique est l'élément structurel fondamental de la biochimie peptidique. C'est la liaison amide entre le carboxyle d'un acide aminé et l'amine du suivant, formée par condensation avec libération d'eau (in vivo via les ribosomes, in vitro via la synthèse peptidique solide), hydrolysable en sens inverse par les enzymes digestives ou les conditions chimiques.
Sa planarité — conséquence d'une résonance partielle entre le carbonyle et l'azote — contraint la géométrie de toute la chaîne polypeptidique et rend possibles les structures secondaires (hélices α, feuillets β) qui définissent la fonction des protéines. C'est sur cette base structurelle que repose toute la pharmacologie peptide — chaque peptide médicament moderne (insuline, sémaglutide, tirzepatide, octréotide) est une chaîne définie de liaisons peptidiques planaires dont la séquence et la conformation déterminent la fonction.
Pour la définition complète du peptide et de la frontière avec la protéine, voir le hub définition. Pour la production industrielle des peptides médicaments, voir synthèse peptidique solide. Pour les peptides alimentaires issus de l'hydrolyse, voir hydrolysat protéique.
Sources
4- 01
- 02
- 03
- 04